| computationally unfolding text procuded by the multicellular automaton Otto B. Wiersma |
27 Sept. 2003 – 15 May. 2004 (last update) |


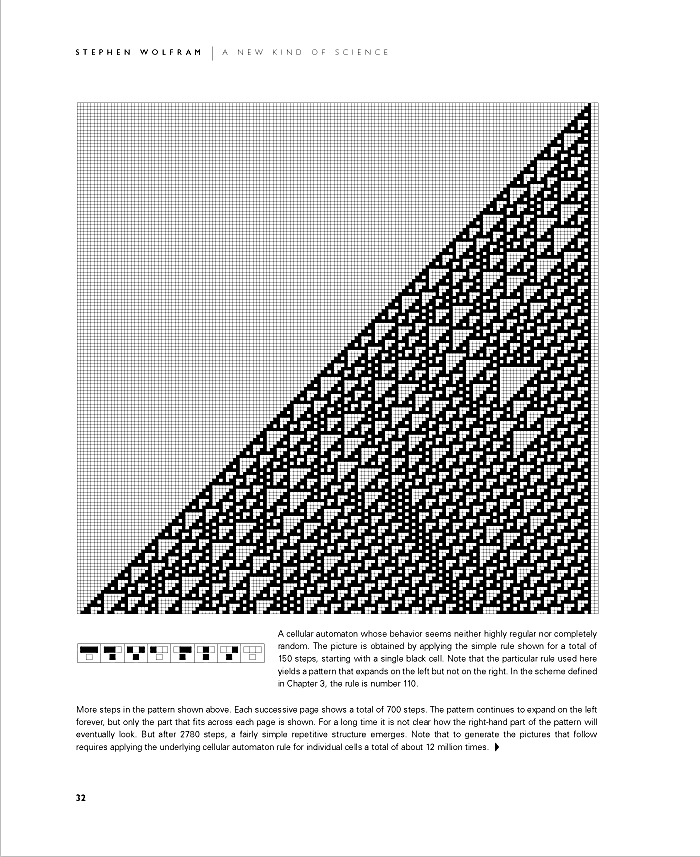
 Behe, M., Darwin’s Black Box. Free Press, 1996
Behe, M., Darwin’s Black Box. Free Press, 1996 Wolfram, S., A New Kind of Science. 2002
Wolfram, S., A New Kind of Science. 2002 Eric Weisstein’s World of Mathematics
Eric Weisstein’s World of Mathematics ISCID Internatinal Society for Complexity, Information and Design Encyclopedia
ISCID Internatinal Society for Complexity, Information and Design Encyclopedia
|
DNA_A = A0000A0A000A = A(5,1,2)+A(4,8,2) DNA_C = 0C0000000CC0 = C/z^2 + C(1,10,2) DNA_G = 00G0G000G000 = G(2,3,2) + G/z^9 DNA_T = 000T00T00000 = T(3,4,2) |
for(i=j=k=1;--j||k;k=j?i%j?k:k-j:(j=i+=2));